Oil-eating Bacteria
Is growth and metabolism of Pseudomonas putida in soil significantly affected by nutrient availability or presence of Hexadecane and are these effects independent?

Abstract
The aim of this experiment was to assess the effects of nutrient content and hexadecane on the growth of Pseudomonas putida, to better understand its environmental requirements for optimal survival, enzyme activity, and catabolism of hydrocarbons in soil. The number of P. putida colonies in 1g of soil was calculated using data from spot plating techniques in various conditions where hexadecane and nutrient availability were manipulated. It was found that nutrient availability significantly affected colonial growth, whereas hexadecane did not. Overall, nutrient supplementation was more beneficial than hexadecane for P. putida survival, although hexadecane may play a role in carbon-limited conditions.
Keywords
Alkane hydroxylase, bioremediation, bioaugmentation, biosurfactants, hydrocarbonoclastic bacteria, rhamnolipids.
Introduction
Pseudomonas putida is a gram-negative, rod-shaped bacterium that is capable of degrading hydrocarbons in oil. This remarkable ability demonstrates potential for use in bioremediation to sustainably and effectively remove oil from contaminated soil (Saadoun, 2002).
P. putida possesses ‘degradative plasmids’ (Chakrabarty, 1974, pp.157–158) which encode enzymes that facilitate the digestion of a range of compounds — including the oxidation of hydrocarbons such as hexadecane (C16H34). The transmissible ability of these plasmids was utilised in 1971 by Prof. Ananda Mohan Chakrabarty, who genetically modified P. putida using plasmid transfer techniques to produce a new, highly efficient strain called ‘multiplasmid hydrocarbon-degrading Pseudomonas’ that was capable of digesting ⅔ of hydrocarbons found in a typical oil spill (Pandey and Arora, 2020). P. putida has since been patented and extensively studied to ensure it is safe, targeted and efficient before it becomes commercially available for use in bioaugmentation processes.
Previous research demonstrates that strains of P. putida utilise hydrocarbons, particularly hexadecane, as carbon and energy sources for growth (Das and Chandran, 2010). Hexadecane as a sole carbon source can induce the synthesis of biosurfactants such as rhamnolipids (Tuleva, Ivanov and Christova, 2014). These amphipathic glycolipids produced by P.putida possess emulsifying properties which assist hydrocarbon degradation and may aid in bacterial motility via changes in cell surface hydrophobicity (Chrzanowski, Ławniczak and Czaczyk, 2011). This suggests hexadecane plays fundamental roles in P. putida population maintenance, as well as their capacity to harness carbon and energy from other hydrocarbon sources.
However, hexadecane alone does not provide P. putida with key nutrients such as nitrogen, phosphorus, and potassium (NPKs), which are necessary for metabolic processes, microbial growth and biofilm maintenance via cell signalling (Zheng et al., 2019; Rapsinski et al., 2024) Although they have been found to thrive in nutrient-limited conditions due to their versatile metabolism (Poblete-Castro et al., 2012) and high rate of NADH regeneration (Blank et al., 2008), NPK supplementation has been found to increase the efficiency of hydrocarbon degradation in soil (Elliott and Fox, 2023). Therefore it is essential to assess the nutritional requirements of P. putida if it is to be manufactured large-scale and used to treat oil-contaminated environments.
Current evidence suggests both hexadecane and nutrient
addition can increase the metabolic activity of
P. putida. To gauge which has a greater influence,
we aim to assess and evaluate the interactive effects of
nutrient and hexadecane availability on
Materials + Methods
PCR and gel electrophoresis was used to confirm the identity of suspected P. putida cultures. 3 diluted cell extracts were aseptically prepared and lysed for PCR: 2 suspected P. putida and 1 was E.coli to allow comparison. 1µl of each diluted extract was transferred to a PCR tube containing 15µl of PCR reaction mix consisting of primers specific to P. putida. A 4th PCR tube containing 1µl of DH20 served as a negative control. After amplification via a PCR machine, samples were mixed with 4µl loading dye. 12µl of a dyed DNA marker was loaded onto a well 1% agarose containing SybrSafe stain, followed by suspected P. putida samples, E.Coli, and the control in separate wells (left to right). Gels were run under an electrical current and digital images were taken via a gel documentation system.
To assess the effects of nutrient and hexadecane contamination on P. putida population growth, we examined 4 samples of sterilised soil inoculated with 1–4x10⁵ CFU/ml P. putida, that had been exposed to different conditions: (NH) soil contaminated with 0.5% (dry weight) hexadecane and treated with a nutrient solution, (H) hexadecane only, (N) nutrient treatment only, and (C) control with no nutrient or hexadecane addition. Water content was adjusted to 50% field capacity and all samples were incubated at 25℃ for 24 h prior to spot plating. Each condition was repeated 6 times to allow for statistical comparison.
Serial dilutions were aseptically set up for each condition. 0.12 g of soil from each sample was aseptically transferred to 4 separate Universal bottles containing 9.98 ml of sterile quarter-strength Ringer solution, and vigorously shaken for 1 minute to produce 10⁻² dilutions for each condition. Following 0.1 ml dilutions (10–3 to 10–6 ) were aseptically prepared in 4 sets of sterile corresponding Eppendorf tubes containing 0.9ml of sterile quarter-strength Ringer solution. 4 nutrient agar spot plates were then prepared for each condition. Spots were marked in a 4x5 grid arrangement. Columns were in order of dilution (10–2 to 10–6) with 4 replicate 5 μl spots per dilution to ensure precision and allow visual comparison of population density. Plates were then incubated at 21℃ for 24h. After incubation, the dilution with the greatest number of visible colonies was selected, and the total count in that dilution was recorded as the number of colonies in 20µl (A). The colony count in 1g of soil (P) was calculated using the following equation:
P = 50( )P for each sample was recorded and analysed using 2-way ANOVA and a post-hoc Tukey HSD, to test for significance as well as interactive effects.
Results



Discussion
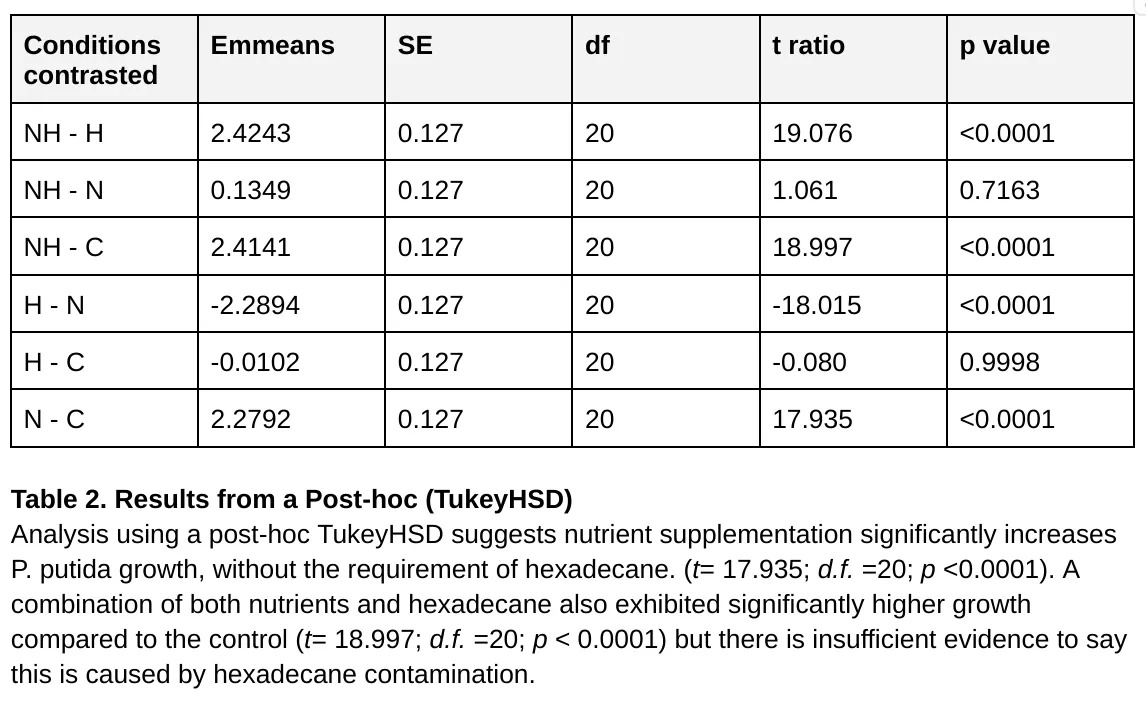
Fig. 2 and statistical analysis confirms that nutrient supplementation has a significant effect on P. putida growth, while the presence of hexadecane alone does not. Furthermore, we found no significant interaction between hexadecane and nutrient presence, suggesting these effects are independent. Overall, nutrient supplementation appears to play a greater role in P. putida growth than hexadecane.
An explanation for this is that nutrients support basic fundamental processes for growth and regular cell function, such as aerobic respiration and protein synthesis (Givskov et al., 1994). This includes the expression at a genetic level, abundance, and ultimate activity of degradative enzymes involved in hexadecane oxidation, such as alkane hydroxylase (AlkB) (Arslan et al., 2014). AlkB catalyses the first step in hexadecane oxidation, requiring iron and nitrogen for activity. This was demonstrated by Mössbauer spectroscopy studies of AlkB’s tertiary structure, identifying a central diiron cluster (Fe-Fe) coordinated by the nitrogen ligands of 8 histidines (Shanklin et al., 1997). This 8-histidine motif facilitates the hydroxylation of various alkane hydrocarbons. Deficiency of nutrients like iron and nitrogen may impair AlkB structure and function, hindering its activity and preventing the bacterium from accessing the metabolic benefits of hexadecane. This highlights how nutrients are limiting factors for P. putida growth and metabolism, particularly hexadecane degradation.
Although the error bars in Fig. 2 suggest that a combination of hexadecane and nutrients results in significantly more growth than nutrients alone, this is not reflected in the 2-way ANOVA or post-hoc test so this statement is unsupported by our data. Contradictory evidence suggests that hexadecane can induce P. putida enzyme activity with adequate nutrients - Alkane hydroxylase activity (U/mL) was increased by the addition of 1% hexadecane by a factor of 8.789 compared to a control culture (Sumarsih et al., 2017).
Alternatively, the growth of P.putida can be affected by biotic factors in the microbial environment. A study on population diversity (Abdel-Megeed, Al-Harbi and Al-Deyab, 2010) found the rate of hexadecane degradation by P. putida and 2 other hydrocarbon- degrading strains (R. erythroplolis and B. thermoleovorans) was higher in combination versus each strain individually. This suggests that P.putidabenefits from other strains to increase efficiency of hydrocarbon degradation, which may result from the diversity in nutrient sources provided by these organisms. Further research should examine the optimal diversity of organisms in P.putida’s microbial community to improve its efficiency in bioremediation.
Despite P.putida being able to thrive on nutrients alone, hexadecane may still play a role as the sole carbon source. This is supported by research that found when provided with a mineral salt medium, P.putida samples inoculated with 1% hexadecane exhibited higher and more sustained colonial growth (OD₆₀₀ ~2.0 by day 9) compared to naphthalene, toluene, and controls without hydrocarbons (OD₆₀₀ ~1.3 by day 4). This suggests a combination of adequate nutrients and hexadecane is optimal for P. putida proliferation. Hexadecane (alkane) may have resulted in more sustained growth than naphthalene/toluene (aromatic hydrocarbons) due to its stability as a linear, saturated hydrocarbon (Chayabutra and Ju, 2000). In contrast, Toluene has been found to suppress growth of P.putida despite providing potential carbon, inducing the synthesis of 22 proteins associated with carbon starvation (Vercellone-Smith and Herson, 1997). This was only when they were used as the sole carbon source, and at high concentrations (267 mg/L). Therefore P.putida is ideal for the bioremediation of crude oil spills, of which stable, saturated hydrocarbons make up the majority (avg. 75.57%) compared to unsaturated hydrocarbons, including aromatics (avg. 14.04%) (Li et al., 2023).
In summary, our research highlights the importance of nutrients as limiting factors for P.putida growth. Regarding the role of hexadecane, it may not be as essential for growth as nutrients, but it provides a crucial function in the bioremediation abilities of P.putida by inducing enzyme activity. Although our data does not support the role of hexadecane, existing literature demonstrates its potential for sustaining growth. Future studies should adopt longer growth periods, as well as assessing interactions between other hydrocarbonoclastic bacteria, to eliminate doubts regarding efficacy and shed light on the potential of P. putida as a tool for bioremediation.
References
Abdel-Megeed, A., Al-Harbi, N. and Al-Deyab, S. (2010). Hexadecane degradation by bacterial strains isolated from contaminated soils. African Journal of Biotechnology, 9(44), pp.7487–7494. doi:10.5897/ajb10.638.
Arslan, M., Afzal, M., Amin, I., Iqbal, S. and Khan, Q.M. (2014). Nutrients Can Enhance the Abundance and Expression of Alkane Hydroxylase CYP153 Gene in the Rhizosphere of Ryegrass Planted in Hydrocarbon-Polluted Soil. PLoS ONE, 9(10), e111208. doi:10.1371/journal.pone.0111208.
Blank, L.M., Georgios Ionidis, Ebert, B.E., Bühler, B. and Schmid, A. (2008). Metabolic response of Pseudomonas putida during redox biocatalysis in the presence of a second octanol phase. FEBS Journal, 275(20), pp.5173–5190. doi:10.1111/j.1742–4658.2008.06648.x.
Chayabutra, C. and Ju, L.-K. (2000). Degradation of n-Hexadecane and Its Metabolites by Pseudomonas aeruginosa under Microaerobic and Anaerobic Denitrifying Conditions. Applied and Environmental Microbiology, 66(2), pp.493–498. doi:10.1128/aem.66.2.493–498.2000.
Chrzanowski, Ł., Ławniczak, Ł. and Czaczyk, K. (2011). Why do microorganisms produce rhamnolipids? World Journal of Microbiology and Biotechnology, 28(2), pp.401–419. doi:10.1007/s11274–011–0854–8.
Elliott, B. and Fox, S. (2023). The Effects of Nutrients on the Efficiency of Motor Oil Bioremediation by Pseudomonas putida. Journal of Student Research, 11(3), n.p. doi:10.47611/jsr.v11i3.1682.
Givskov, M., Eberl, L., Möller, S., Poulsen, L. and Molin, S. (1994). Responses to nutrient starvation in Pseudomonas putida KT2442: analysis of general cross-protection, cell shape, and macromolecular content. Journal of Bacteriology, 176(1), pp.7–14. doi:10.1128/jb.176.1.7–14.1994.
Li, Y., Hou, D., Cheng, X., Han, X. and Niu, C. (2023). Geochemical characteristics and significance of aromatic hydrocarbons in crude oil from the East Fukang Sag, Junggar Basin, NW China. Frontiers in Earth Science, 10, n.p. doi:10.3389/feart.2022.1031367.
Pandey, P. and Arora, N.K. (2020). Prof. Ananda Mohan Chakrabarty: The Superbug Superhero! Environmental Sustainability, 3, pp.333–335. doi:10.1007/s42398–020–00117-x.
Poblete-Castro, I., Escapa, I.F., Jäger, C., Puchalka, J., Chi Lam, C., Schomburg, D., Prieto, M. and Martins dos Santos, V.A. (2012). The metabolic response of P. putida KT2442 producing high levels of polyhydroxyalkanoate under single- and multiple-nutrient-limited growth: Highlights from a multi-level omics approach. Microbial Cell Factories, 11(34), n.p. doi:10.1186/1475–2859–11–34.
Rapsinski, G.J., Michaels, L.A., Hill, M., Yarrington, K.D., Haas, A.L., D’Amico, E.J., Armbruster, C.R., Zemke, A., Limoli, D. and Bomberger, J.M. (2024). Pseudomonas aeruginosa senses and responds to epithelial potassium flux via Kdp operon to promote biofilm. PLoS Pathog, [online] 20(5), e1011453. doi:10.1371/journal.ppat.1011453.
Saadoun, I. (2002). Isolation and characterization of bacteria from crude petroleum oil contaminated soil and their potential to degrade diesel fuel. Journal of Basic Microbiology, [online] 42(6), pp.420–428. doi:10.1002/1521–4028(200212)42:6%3C420::aid-jobm420%3E3.0.co;2-w.
Shanklin, J., Achim, C., Schmidt, H., Fox, B.G. and Munck, E. (1997). Mossbauer studies of alkane ω-hydroxylase: Evidence for a diiron cluster in an integral-membrane enzyme. Proceedings of the National Academy of Sciences, 94(7), pp.2981–2986. doi:10.1073/pnas.94.7.2981.
Sumarsih, S., Ni’matuzahroh, N., Fatimah, F. and Puspitasari, M. (2017). Effect of aliphatic and aromatic hydrocarbons on the oxygenase production from hydrocarbonoclastic bacteria. Journal of Chemical Technology and Metallurgy, 52, pp.1062–1069. Available at: https://www.researchgate.net/publication/
321127630_Effect_of_aliphatic_and_aromatic_hydrocarbons_on_the_oxygenase_production_from
_hydrocarbonoclastic_bacteria [Accessed 6 May 2025].
Tuleva, B.K., Ivanov, G.R. and Christova, N.E. (2014). Biosurfactant Production By A New Pseudomonas Putida Strain. Zeitschrift für Naturforschung C, 57(3–4), pp.356–360. doi:10.1515/znc-2002–3–426.
Vercellone-Smith, P. and Herson, D.S. (1997). Toluene Elicits a Carbon Starvation Response in Pseudomonas putida mt-2 Containing the TOL Plasmid pWW0. Applied and environmental microbiology, 63(5), pp.1925–1932. doi:10.1128/aem.63.5.1925–1932.1997.
Zheng, L., Ren, M., Xie, E., Ding, A., Liu, Y., Deng, S. and Zhang, D. (2019). Roles of Phosphorus Sources in Microbial Community Assembly for the Removal of Organic Matters and Ammonia in Activated Sludge. Frontiers in Microbiology, 10(1023). n.p. doi:10.3389/fmicb.2019.01023.